
Desert Woodrat
Distribution, Abundance, and Seasonality
The desert woodrat occurs in California in 2 disjunct areas. It is found in northeastern California in Great Basin areas of eastern Modoc Co. to southeastern Lassen Co. Inhabits virtually all of southern California, with range extending northward along the coast to Monterey Co., and along the Coast Range to San Francisco Bay. In southeastern California, found from Inyo Co. south throughout the Mojave Desert and from north-central Tulare Co. south through the Tehachapi and San Bernardino Mts. Common to abundant in Joshua tree, pinyon-juniper, mixed and chamise-redshank chaparral, sagebrush, and most desert habitats. Also found in a variety of other habitats. Most abundant in rocky areas with Joshua trees. Elevational range from sea level to 2600 m (8500 ft). Northern and elevational distribution may be limited by temperature (Lee 1963, MacMillen 1964).
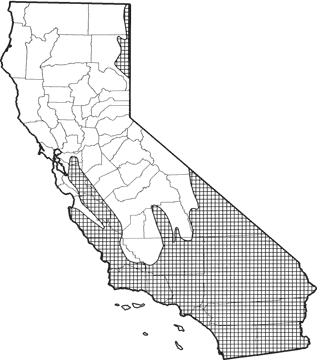
Range Map
Specific Habitat Requirements
Feeding: Eats buds, fruits, seeds, bark, leaves, and young shoots of many plant species. In coastal scrub, prefers live oak, chamise, and buckwheat as food plants (Meserve 1974). Creosote, cholla, and prickly-pear are eaten in the Mojave Desert (MacMillen 1964, Cameron and Rainey 1972). In juniper/sagebrush habitats, ate Mormon-tea, rattlesnake weed, mustard, sagebrush, and buckwheat (Stones and Hayward 1968).
Cover: Houses are constructed with twigs, sticks, cactus parts, rocks, depending on availability of building materials. The house usually is built against a rock crevice, at the base of creosote or cactus, or in the lower branches of trees. Rock crevices appear preferred where available, but woodrats generally adapt to virtually any situation. Houses are used for nesting, food caching, and predator escape.
Reproduction: Nests of dried vegetation, usually fibrous grass parts or shredded stems, are located within the stick house. Suitable nesting sites or nesting materials may limit distribution.
Water: Largely dependent upon prickly pear for water balance in desert habitats, although can be sustained on creosote year-round (Lee 1963, MacMillen 1964).
Pattern: Moderate to dense canopies preferred. Desert woodrats are particularly abundant in rock outcrops and rocky cliffs and slopes (Hall 1946, Miller and Stebbins 1964).
Species Life History
Activity Patterns: Active yearlong. Mainly nocturnal, but also crepuscular and occasionally diurnal (Stones and Hayward 1968, Miller and Stebbins 1964).
Seasonal Movements / Migration: None.
Home Range: In coastal sage scrub, home range was about 0.04 to 0.2 ha (0.10 to 0.5 ac) (MacMillen 1964, Bleich and Schwartz 1975). Average linear movements in same habitat were about 14 m (46 ft)/night. In sagebrush-juniper habitat, males moved 80 m (262 ft)/night, and females 45 m (147 ft) (Stones and Hayward 1968). In coastal sage habitat, density averaged 3.5 to 12.3/ha (1.4 to 4.9/ac) in one study (MacMillen 1964) and 30/ha (12/ac) in another (Bleich and Schwartz 1975). In jumping cholla cactus habitat, density averaged 38/ha (15/ac) (Brown et al. 1972). In sagebrush-juniper habitat densities averaged 2.8/ha (1.1/ac) (Stones and Hayward 1968).
Territory: Aggressively solitary. Territory probably equals home range. Woodrats may defend succulent plants (water sources) against other species, and perhaps prevent other species from obtaining water during droughts (MacMillen 1964).
Reproduction: Breeds from October to May, depending on the habitat. Nesting is solitary. Gestation period is 30-36 days (Egoscue 1957). Litter size averages 2.7 (range 1-5) (Egoscue 1957, MacMillen 1964). Polyestrous in lab, but probably breeds once/yr in wild (Egoscue 1957). Weaning is at 27-40 days (Egoscue 1957, Cameron 1973). Females may breed at 2-3 mo of age.
Niche: The desert woodrat is a moderate-sized folivore/granivore. Competitors include cricetid and heteromyid rodents. Woodrat houses provide shelter for a variety of small vertebrates. Predators include snakes, owls, and predatory mammals. This woodrat is commonly parasitized by bot fly larvae.
Sources & References
California Department of Fish and Game, 1999.
California's Wildlife, Sacramento, CA.
Written by: P. Brylski, reviewed by: H. Shellhammer, edited by: R. Duke
Bleich, V. C., and O. A. Schwartz. 1975. Observations on the home range of the desert woodrat, Neotoma lepida intermedia. J. Mammal. 56:518-519. Brown, J. H., G. A. Lieberman, and W. F. Dengler. 1972. Woodrats and cholla: dependence of a small mammal population on the density of cacti. Ecology 53:310-313. Cameron, G. N. 1973. Effect of litter size on postnatal growth and survival in the desert woodrat. J. Mammal. 54:251-256. Cameron, G. N., and D. G. Rainey. 1972. Habitat utilization by Neotoma lepida in the Mohave Desert. J. Mammal. 53:251-266. Egoscue, H. J. 1957. The desert woodrat: a laboratory colony. J. Mammal. 38:472-481. Hall, E. R. 1946. Mammals of Nevada. Univ. California Press, Berkeley. 710pp. Lee, A. K. 1963. The adaptations to arid environments in woodrats of the genus Neotoma. Univ. Calif. Publ. Zool. 64:57-96. MacMillen, R. E. 1964. Population ecology, water relations and social behavior of a southern California semidesert rodent fauna. Univ. Calif. Publ. Zool. 71:1-59. Meserve, P. L. 1974. Ecological relationships of two sympatric woodrats in a California coastal sage scrub communty. J. Mammal. 55:442-447. Miller, A. H., and R. C. Stebbins. 1964. The lives of desert animals in Joshua Tree National Monument. Univ. California Press, Berkeley. 452pp. Stones, R. C., and C. L. Hayward. 1968. Natural history of the desert woodrat, Neotoma lepida. Am. Midl. Nat. 80:458-476.
California Animal Facts | California's Wildlife